2. Cytologie et biochimie
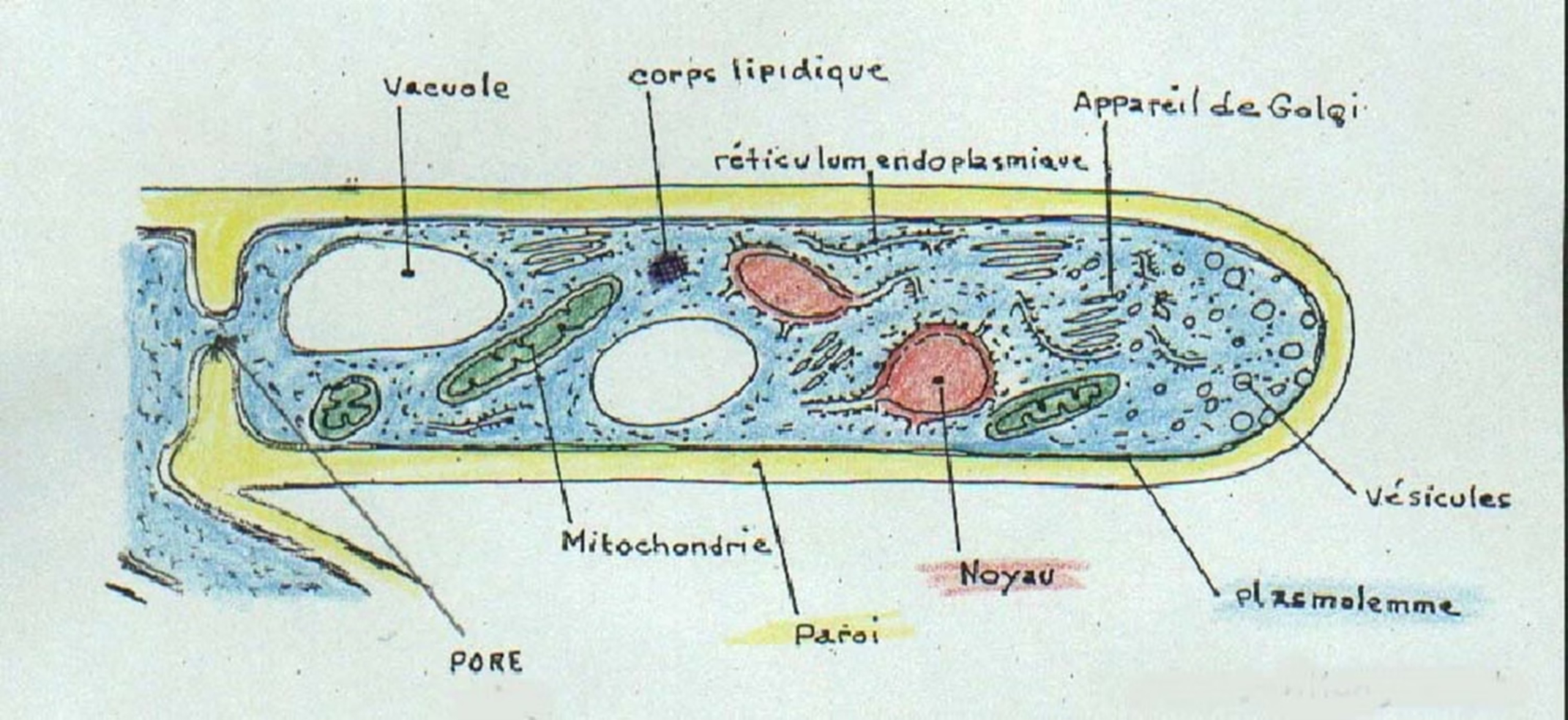
La cellule fongique est formée par le noyau (certaines structures en ont un, d’autres deux ou plus), les mitochondries, l’appareil de Golgi, le reticulum endoplasmique, les ribosomes, les vacuoles de réserves (contenant du glycogène), le cytosquelette (filaments d’actine et de tubuline), la membrane cytoplasmique (plasmalemme) et la paroi.
1. Le Noyau:
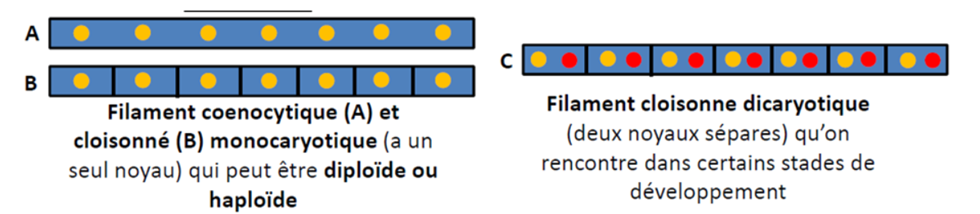
Différents types d’organisations nucléaires sont rencontrés à divers stades du cycle de développement : haploïde, diploïde et dicaryotique.
2. Les réserves
Les réserves chez les champignons sont sous forme de glycogène (comme dans les hépatocytes humains) (chez les plantes amidon) ou de gouttelettes lipidiques.
3. Vacuole: La vacuole constitue un important stock enzymatique.
4. Les septa
Ce sont formés par une extension localisée de la couche interne de la paroi de l’hyphe vers le centre de celle-ci, à la manière d’un diaphragme d’appareil photo qui se ferme.
Dans de nombreux cas, les septa sont incomplets et possèdent un ou plusieurs pores qui restent ouverts pendant la majorité de la période de croissance de l’hyphe, permettant le libre passage du cytoplasme entre les articles adjacents.
Le rôle des septa
Les septa ont plusieurs fonctions :
* ils servent à renforcer et stabiliser ces longs tubes que sont les hyphes .
* ils servent aussi de protection au mycélium. En cas de brisure d’une hyphe, ils se ferment et isolent l’article brisé du reste de l’hyphe, protégeant celle-ci du milieu extérieur.
*Enfin, en isolant les articles du reste de l’hyphe, ils permettent la différenciation et la spécialisation des articles et leur transformation en hyphes modifiées.
Les différents types de septa.
En général, les zygomycètes n’ont pas de septa, mais parfois des septa complètement clos peuvent se former pour isoler de vieilles sections ou des sections endommagées du mycélium. (Fig. 2-A).
Les ascomycètes ont des septa perforés dont le pore peut être ouvert ou fermé par un corps membraneux composé de protéines appelé « corps de Woronin » (Fig. 2-B).
Le pore des ascomycètes est relativement grand (0,05 à 0,5 µm) et permet le passage des organites cytoplasmiques et même des noyaux. Après qu’un pore a été obstrué par un corps de Woronin, il ne retrouve jamais sa perméabilité.
La croissance de l’hyphe peut quand même se poursuivre à partir d’un nouvel apex formé derrière l’article endommagé ou d’un nouvel apex qui croît à l’intérieur de l’article endommagé à partir du septum. Les septa des ascomycètes
sont aussi caractérisés par l’absence de parenthésome.
Mais le plus complexe des septa est celui que possèdent les basidiomycètes (Fig. 2-C).
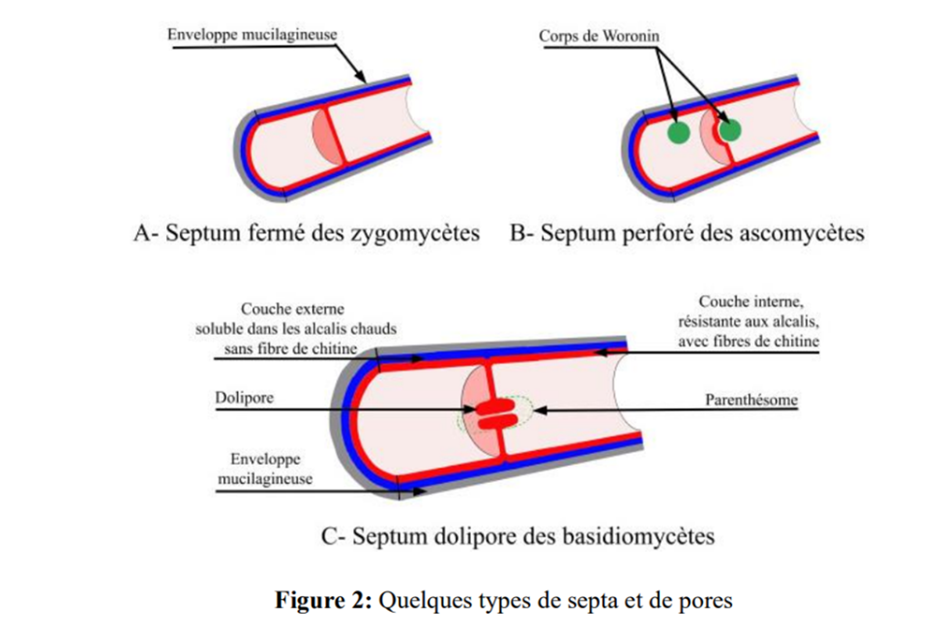
Les basidiomycètes ont des septa qui divisent l’hyphe en articles. Ces septa possèdent un pore central relativement étroit (100-150 nm) ayant la forme d’un beignet, appelé dolipore (dolium = tonneau), qui peut se fermer complètement ou s’ouvrir et servir de filtre. Au microscope optique, le dolipore apparaît comme un petit bouton central au septum.
Dans des conditions favorables, le pore lui-même peut-être observé (Figure 3). Grâce à ce dolipore, il existe une continuité entre les articles qui permet au cytoplasme, aux noyaux et aux mitochondries de migrer d’un article à l’autre le long de l’hyphe
À une certaine distance de chaque côté du dolipore se trouve une structure, la plupart du temps perforée, qui ressemble à une paire de parenthèses, d’où son nom, parenthésome. Le parenthésome c'est une membrane hémisphérique perforée provient du réticulum endoplasmique et serait une structure ancestrale au dolipore; il assurerait la continuité cytoplasmique tout en bloquant le passage aux organites majeurs. Les dolipores et les parenthésomes agissent ensemble comme un tamis dans le cytoplasme.