VI- La pollinisation, la fécondation et l'embryogenèse
1- La pollinisation : Pour qu'il y ait pollinisation, il faut que le pollen arrive jusqu'au carpelle, pour cela, il s'offre différent choix si l'on peut dire :
1-1- Autopollinisation (autogamie) : le pollen est transporté sur le stigmate de la même fleur. Cela est rare (autoincompatibilité) pour éviter les conséquences néfastes de la dégénérescence liée à l’endogamie.
1-2- Pollinisation croisée (allogamie) : le pollen est transporté sur le stigmate d'une autre fleur. C'est évidemment le plus représenté des transports.
Ce transport peut se faire par le vent (Pollinisation Anémophile), ou par les animaux (Pollinisation zoophile : les insectes (Entomophile), les oiseaux (Ornithophile) et les chauves-souris (Cheiroptérophile)). La pollinisation par l’eau (hydrophile) est rare.

1-3- Les mécanismes permettant d’évité l’autopollinisation :
a) Séparation spatiale et temporelle des gamètes mâles et femelles :
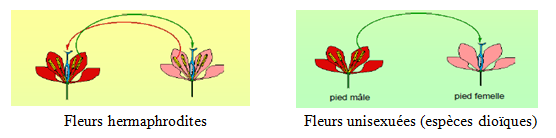
- Spatiale : dioécie (espèce dioïque) ou monoécie (espèce monoïque)
- Temporelle : protogynie (stigmate réceptif ou mûr avant l’émission du pollen dans une fleur hermaphrodites ou monoïque) ou protandrie (émission du pollen quand les stigmates ne sont pas encore réceptifs)
b) Autoincompatibilité : impossibilité pour une plante hermaphrodite de produire des zygotes avec son propre pollen. Cette autoincompatibilité est contrôlée génétiquement et peut être :
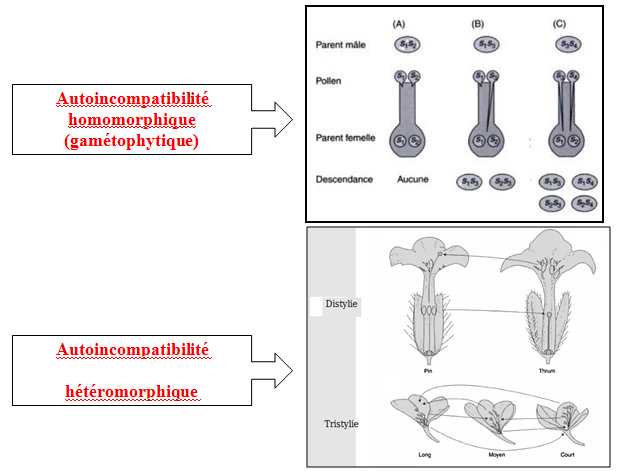
- Homomorphique (morphologie homogène) : gamétophytique ( le génotype du pollen détermine la pollinisation) ou sporophytique (le génotype du sporophyte détermine le sort des grains de pollen).
- Hétéromorphique (morphologie hétérogène des styles et des étamines) : distylie (Pin et Thrum) ou tristylie (Long, Moyen et Court).
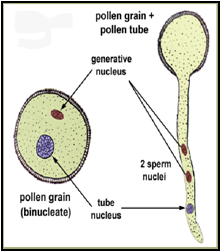
1-4- La germination du grain de pollen : Des phénomènes de reconnaissance au niveau moléculaire assurent une sélection des pollens (filtrage génétique). Cette reconnaissance se fait au niveau de protéines du tube pollinique (papille stigmatique). Dans le cas où le grain de pollen est compatible, il pourra germer. Au départ, il contient deux noyaux :
-Le noyau génératif, aussi appelé spermatogène, contient le matériel génétique. Il progressera de concert dans le tube pollinique et pendant son parcours il se scindera en deux noyaux spermatiques qui auront chacun leur rôle lors de la double fécondation de l'ovule.
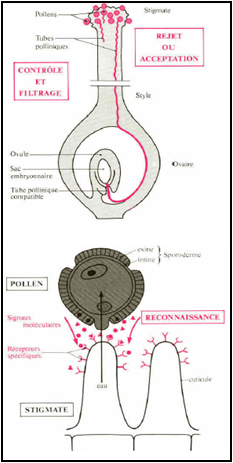
Le grain de pollen se développera et passera entre les cellules du style et atteindra l'ovule soit par le micropyle (Porogamie) soit par la chalaze (Chalazogamie). La pénétration du tube pollinique dans le style se fait par la sécrétion d’enzymes digestives. Le guidage du tube pollinique dans le pistil peut s’expliquer par un contrôle mécanique dû à l’architecture et la longueur du pistil de l’espèce receveuse. Les tubes polliniques incompatibles rencontrent des obstacles dans leur course ce qui limite la réussite de la fécondation, alors que ceux compatibles sont attirés par des signaux chimiques (micropyle, synergides…). Le tube pollinique pour arriver à l'ovule passe par une des deux synergides et passe dans l'appareil filiforme de la synergide, puis les gamètes mâles sont libérés.
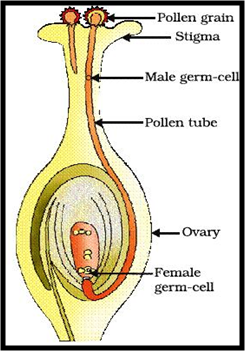
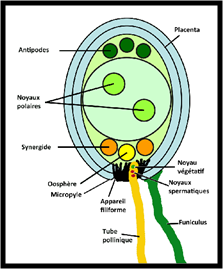
2- La fécondation : Après le passage du tube pollinique dans l’appareil filiforme des deux synergides et la libération des deux gamètes mâles, l'un et l'autre interviennent dans la fécondation du même sac embryonnaire, c'est pourquoi l'on parle de double fécondation. L'un d'entre eux féconde l'oosphère, l'autre s'unit aux deux noyaux « polaires » de la cellule centrale du sac embryonnaire. La première fécondation aboutit à la formation d'un zygote diploïde (2n), qui évoluera en embryon ; la deuxième donne un zygote triploïde (3n) qui évoluera en albumen. Après cette double fécondation, le pistil (carpelle) reprend sa croissance et évolue en fruit tandis que l’ovule se transforme en graine.
3- Le développement du zygote (Embryogenèse)
La double fécondation aboutit à la formation d'un zygote diploïde (2n), qui évoluera en embryon et un zygote triploïde (3n) qui évoluera en albumen. Ainsi l’ovule fécondé évolue pour former la graine composée de :
A) Les téguments : Après la fécondation, le micropyle se ferme, les téguments de l'ovule deviennent ceux de la graine : le tégument externe devient dure et épais, c’est le testa, il peut être coloré, présenter des poils …, et le tégument interne devient mince et se nomme le tegmen. Remarque : il peut y avoir des formations externes supplémentaires comme des excroissances des téguments externe (ex : la caroncule)
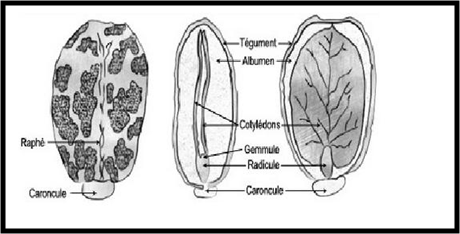
B) L’embryon : Le zygote principal (2n) subit une suite de segmentations rigoureusement orientées. Par divisions transversales répétées, une file de cellules est édifiée. On distingue, du côté du micropyle, le suspenseur et, vers l’intérieur, une cellule terminale qui, par divisions longitudinales puis périclines, permet la mise en place des principaux organes : Ebauche des cotylédons; Ebauche de la tigelle ou l’hypocotyle se prolongeant de la radicule; Procambium : qui formera les premiers faisceaux vasculaires; Protoderme : future épiderme; Ebauche de l’épicotyle ou la gemmule.
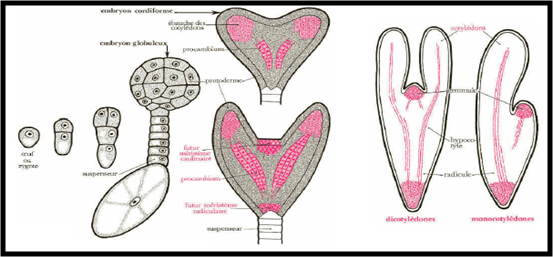
C)- L’albumen :
Le zygote accessoire (3n) se divise activement ; les noyaux produits restent tous d’abords libres. La cellularisation débute ensuite par la périphérie de ce qui fut le sac embryonnaire et progresse vers le centre. Les cellules se chargent de réserves formant un tissu nourricier, l'albumen, autour de l'embryon issu du zygote principal.
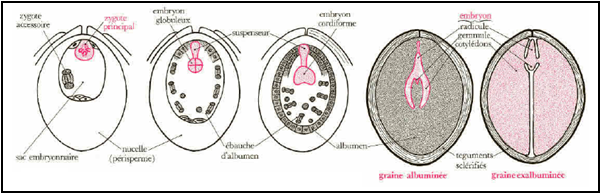
- Graines albuminées (ex. : céréales). L'albumen persiste et sert de réserve alimentaire pour le développement futur de l'embryon au cours de la germination.
- Graines exalbuminées (ex. : légumineuses) L’albumen est entièrement consommé par l'embryon au cours du développement de celui-ci, comme tout ce qui reste du nucelle lui-même, voire encore le tégument interne. Les matières de réserve sont alors stockées dans les cotylédons.
- Graines périspermées (ex : poivre) Il n'est pas rare que persiste, autour de l'albumen à maturité, une portion périphérique du nucelle, le périsperme, qui peut parfois remplacer l'albumen.
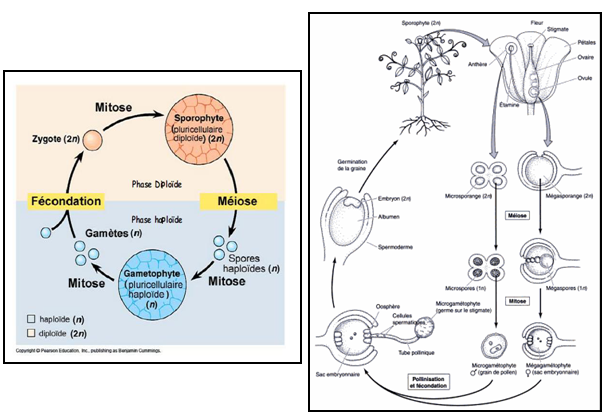
4- Le cycle de développement
Toutes les cellules de l’appareil végétatif renferment le nombre normal de chromosomes de l’espèce, soit 2n chromosomes. On dit que les noyaux sont diploïdes. Mais nous avons vu que les cellules mères des grains de pollen et les cellules mères du sac embryonnaire subissent une division particulière qui réduit de moitié le nombre des chromosomes à n chromosome de leur noyau qui devient alors haploïde. Puis à lieu la fécondation, et deux noyaux haploïdes donnent le noyau diploïde de l’oeuf fécondé (Zygote). Nous pouvons donc distinguer deux phases dans la vie de la plante : une phase diploïde où les cellules ont 2n chromosomes, et une phase haploïde avec des cellules à n chromosomes ; elles sont séparées par deux phénomènes importants : la réduction chromatique (méiose) et la fécondation. Nous remarquons qu’ici, la phase diploïde est de loin la plus importante..